
All around Bioenergetics
Peter Mitchell's Little Grey Book


MPTP, mitochondrial Complex I, and William Gibson's Necromancer
"Peter's good as dead anyway," Molly said. "In another twelve
hours, he'll start to freeze up. Won't be able to move, his eyes
is all."
"Why?" Case turned to her.
"I poisoned his sh*t for him," she said. "Condition's like
Parkinson's disease, sort of."
3Jane nodded. "Yes. We ran the usual medical scan, before he was
admitted." She touched the ball in a certain way and it sprang away from
Molly's hands. "Selective destruction of the cells of the substantia
nigra. Signs of the formation of a Lewy body. He sweats a great deal, in his
sleep."
"Ali," Molly said, ten blades glittering, exposed for an instant. She
tugged the blanket away from her legs, revealing the inflated cast.
"It's the meperidine. I had Ali make me up a custom batch. Speeded up
the reaction times with higher temperatures. N-methyl-4-phenyl-1236," she
sang, like a child reciting the steps of a sidewalk game,
"tetra-hydro-pyridene."
"A hotshot," Case said.
"Yeah," Molly said, "a real slow hotshot."
"That's appalling," 3Jane said, and giggled.
from "Necromancer" by William Gibson, 1984 (view it here )
Apparently american Sci-Fiction writer William Gibson was at the front of biochemical research of his time. MPTP or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine was first manufactured in 1947 and used mostly as a starting material for the synthesis of other organic substances. It was even tested as a possible treatment for Parkinson’s Disease (PD). In 1982 the effects of MPTP were discovered in a long chain of events when the particular batch of illegal synthetic meperidine drug (sold under the name of China White) induced Parkinson’s-like symptoms in several young drug users in Santa Clara County, California. Almost overnight they became “frozen” in a PD-like state [Langston at al., 1983 ]. Doctors were puzzled by these cases - the patients were too young and deteriorating too rapidly to suffer from clinical PD. After analysis of paraphernalia used by the drug addicts, consideration of their symptoms and autopsy results it turned out that MPTP was the key factor selectively toxic to the substantia nigra [Langston at al., 1983 ]. The substantia nigra is an area of the midbrain responsible for movement control and death of dopaminergic neurons in that area is the cause of Parkinson’s disease resulting in tremor, stiffness and slowness of motor movements.
However, even as early as 1976, a young chemistry research student from Maryland, Barry Kidston, synthesised meperidine incorrectly which led instead to the production of the toxin MPTP. After testing his product on rats (rodents turned out to be much less sensitive to MPTP), he injected himself with his own product and within three days developed symptoms of Parkinsonism [Davis et al., 1979].
It has been shown that once taken up in the brain, MPTP is converted by monoamineoxidase (MOA-B) in glial cells into another compound, MPP+, which is transported by the dopamine transporter of surrounding neurons and inhibits the important cellular organelle, the mitochondria (Figure 1). MPP+ is very weak inhibitor of mitochondrial Complex I but as a lipophilic cation (positively charged) it is accumulated into the mitochondrial matrix in accordance with the mitochondrial membrane potential (negative inside). Even if present at a nanomolar range outside the cells, MPP+ concentration in mitochondria could reach millimolar levels - enough to inhibit the enzyme. Interestingly, MPTP resulted in death even in flatworms, the most primitive species with a central nervous system, but not in rodents which lack the specific dopamine transporter.
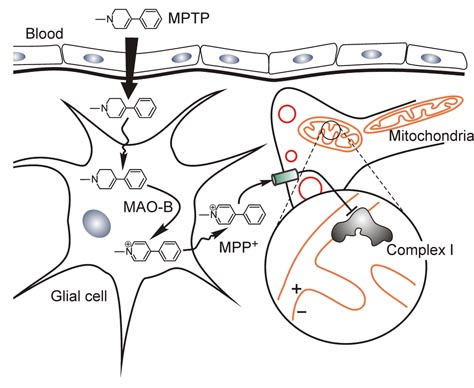
Figure 1. MPTP can cross the blood–brain barrier and is metabolized to MPP+ (1-methyl-4-phenylpyridinium) with the assistance of the enzyme monoamine oxidase B (MAO-B) in glial cells. MPP+ is then taken up by into neuronal cells dopamine transporter (shown green) for which it has high affinity. Once inside neurons, MPP+ is concentrated by an active electrophoretic transport within the mitochondria, where it impairs mitochondrial oxidative phosphorylation by inhibiting complex I of the respiratory chain.
Despite of intensive research of Parkinson disease several questions remains to be answered:
(i) Why are dopaminergic neurons so sensitive especially to Complex I inhibition?
(ii) What is the cause and the mechanism of selective inhibition of Complex I in PD ?
(iii) Why is activity of complex I lower not only in neurons of PD-patients but also in platelets from the blood?
(iv) What is the relative role of possible oxidative stress, mitochondrial DNA damage or energy-deficit in the development of PD?
You could read further about that fascinating story in the The Case of the Frozen Addicts book by J. William Langston
References
Langston JW, Ballard P, Tetrud JW, Irwin I. (1983) Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science. 219, 979
Davis, G.C., Williams, A.C., Markey, S.P., Ebert, M.H., Caine, E.D., Reichert, C.M., Kopin, I.J., (1979) Chronic Parkinsonism secondary to intravenous injection of meperidine analogues. Psychiatry Res. 1, 249
Fishing, Thomas Mayne Reid and Complex I inhibitor rotenone

From The Forest Exiles by Mayne Reid 1854 (can be found here )
As mentioned by Thomas Reide, fishing by water poisoning using roots of some tropical plants was common in Amazonia (in fact it still is, see video). Specific compounds called rotenoids are responsible for poisoning of fish and can be found almost exclusively among legumes in the family Fabaceae. Rotenone (Fig 2), the best known member of the group, was first extracted by French botanist Emmanuel Geoffroy in 1895 under the name of “nicouline” from a native plant of French Guiana called “Robinia nicou” (now Lonchocarpus nicou) [Geoffroy 1895].
The wide distribution of rotenone-containing plants explains why this toxicant has been used for a long time as a fish poison by people of many different cultures. The procedure was first documented among natives of the East Indies and South-East who crushed the roots of tuba plant (liana Derris elliptica), sometimes along with small shrimps, mixed it with human excrement and poisoned water in small rivers with this mixture [Rumphius 1747]. The employment of derris extract as an insecticide by gardeners in the East Indies has long been a common practice. The use of Derris elliptica extracts against leaf eating caterpillars during nutmeg cultivation was also reported [Oxley 1848].
In 1902 Kazuo Nagai in Japan extracted a crystalline substance from the roots of Derris chinensis, which he called rotenone from "roh-ten" which was the name of the plant in Taiwanese [Nagai 1902]. In 1929 rotenone was isolated from the roots of Peruvian plant Lonchocarpus sp., locally known as barbasco or cube. Lonchocarpus plants even became an export product as an insecticide due to their high content of rotenoids. In the beginning of the XX century rotenone was used as a remedial agent for combating mites, fleas and body lice on animals and as a killing agent for house flies and roaches [Ambros&Haag 1936] (see the 40's advertisment).
Rotenoids are a special group of Complex I inhibitors. When rotenone is introduced to the water by mashing the part of the plants, it lyses fish gills and half-paralysed fish are forced to swim at the water surface where they can be easily caught. In early studies it was found that rotenone inhibits oxygen consumption in fish gill filaments, liver tissue slices and isolated liver mitochondria [Lidhal&Oberg, 1961]. It was found to be a powerful inhibitor of the mitochondrial aerobic oxidation of substrates linked to NADH without affecting succinate oxidation [Ernster et al, 1963]. Later, the target of rotenone was identified to be mitochondrial Complex I and it was used widely by researchers in the mitochondrial field. As with many hydrophobic substances inhibiting the enzyme, the rotenone molecule structurally resembles Complex I substrate ubiquinone in one of its folded forms (Fig 2). It is generally accepted that rotenone blocks the electron transfer between the terminal iron-sulphur cluster N2 of the enzyme and the ubiquinone pool. Most likely rotenone binds close to the ubiquinone-binding site of the enzyme preventing entrance of a substrate. Many of the common hydrophobic inhibitors of Complex I are found to share a common binding domain in the vicinity of ubiquinone binding site [Okun et al., 1999]. Due to the fact that interaction between the enzyme and rotenone is mediated through the membrane phase, its kinetic model cannot be described as classical competitive inhibition with ubiquinone. The kinetics of rotenone binding is not trivial and is characterised in terms of tightly-bound reversible inhibitor binding to two non-interacting binding sites with different affinities [Grivennikova et al, 2003].
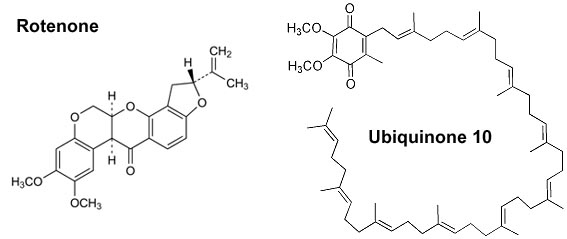
Figure 2. Structure of rotenone (left) and ubiquinone UQ-10 (right) molecule
References

Ambrose A, Haag H (1936) Toxicological study of Derris. Ind. Eng. Chem. 28, 815
Ernster L, Dallner G, Azzone G 1963 Differential effects of rotenone and amytal on mitochondrial electron and energy transfer. J. Biol. Chem. 238, 1124
Geoffrey, E. 1895. Contribution à l'étude du Robinia Nicou Aublet, au point de vue botanique, chimique et physiologique”. Ann. Inst. Colon. Marseille 2, 1
Grivennikova VG, Roth R, Zakharova NV, Hägerhäll C, Vinogradov AD. (2003) The mitochondrial and prokaryotic proton-translocating NADH:ubiquinone oxidoreductases: similarities and dissimilarities of the quinone-junction sites. Biochim Biophys Acta. 1607, 79
Lindahl P.E., Oberg K.E.(1961) The effect of rotenone on respiration and its point of attack. Exp.Cell Res. 23, 228
Nagai, K. (1902). Research on poisonous principles of rohton J. Tokyo Chemi. Soc. 23, 744
Okun JG, Lümmen P, BrandtU.(1999) Three classes of inhibitors share a common binding domain in mitochondrial complex I (NADH:ubiquinone oxidoreductase. J Biol Chem. 274, 2625
Oxley T (1848) Some account of the nutmeg and its cultivation J. Indian Archipelago and East Asia, 10, 641
Rumphius GE (1747) Hermarium Amboinense, Vol 5, Amsterdam, 492.